Наряду с популяцией в генетике существует понятие «чистая линия» - это потомство, полученное только от одного родителя и имеющее с ним полное сходство по генотипу
Чистые линии могут быть созданы в растениеводстве у самоопыляющихся растений. В отличие от популяций они характеризуются полной гомозиготностью. Вследствие полной гомозиготности отбор в чистой линии невозможен, так как все особи, входящие в нее, имеют идентичный набор генов. Высокогомозиготных линейных мышей, крыс и других лабораторных животных создают в целях проведения различных экспериментов, например для проверки на мутагенность тех или иных препаратов, оценки вакцин и т. д.
Популяция состоит из животных разных генотипов. Эффективность отбора в ней зависит от степени генетической изменчивости - соотношения доминантных и рецессивных генов. Харди и Вайнберг провели математический анализ распределения генов в больших популяциях, где нет отбора, мутаций и смешивания популяций. Они установили, что такая популяция находится в состоянии равновесия по соотношению генотипов, что определяется формулой
р2АА + + IpqAa + q}aa =1,
где р - частота доминантного гена А,
q - частота его рецессивного аллеля а.
В соответствии с этим был сформулирован закон, или правило, Харди - Вайнберга, согласно которому при отсутствии факторов, изменяющих частоты генов, популяции при любом соотношении аллелей от поколения к поколению сохраняют эти частоты аллелей постоянными. Несмотря на известные ограничения, по формуле Харди - Вайнберга можно рассчитать структуру популяции и определить частоты гетерозигот (например, по летальным или сублетальным генам, зная частоты гомозигот по рецессивным признакам и частоты особей с доминантным признаком), проанализировать сдвиги в генных частотах по конкретным признакам в результате отбора, мутаций и других факторов.
Популяция находится в равновесии только тогда, когда в ней не происходит отбора. При выбраковке же отдельных животных в такой популяции изменяется соотношение гамет, что влияет на генетическую структуру следующего поколения. Однако К. Пирсон показал, что, как только возникает состояние панмиксии (свободное скрещивание), соотношение генотипов и фенотипов в популяции в следующем поколении возвращается к тому, которое соответствует формуле Харда - Вайнберга, но уже при другом их соотношении. Скрещивание, восстанавливающее соотношение генотипов в популяции, в соответствии с формулой Харди - Вайнберга получило название стабилизирующего. Из этого следует вывод: при использовании в популяции случайных, неотобранных производителей или маток наблюдается стабилизация признаков продуктивности на одном уровне, и повышение продуктивности животных в такой ситуации невозможно. Точно так же при отсутствии браковки гетерозиготных носителей рецессивных аномалий частота проявления аномальных животных в популяции остается неизменной.
В популяциях сельскохозяйственных животных постоянно изменяются частоты генов, что можно наблюдать при анализе смежных поколений. Такие изменения составляют суть генетической эволюции. Основные факторы эволюции: мутации, естественный и искусственный отбор, миграции, дрейф генов.
Одна из основных причин генетической изменчивости в популяции - мутации. Спонтанные мутации каждого гена происходят с низкой частотой, однако общая частота мутаций всех генов, которые содержат популяции, очень велика. Мутации, возникающие в половых клетках родительского поколения, приводят к изменению генетической структуры у потомства. В популяции постоянной численности в отсутствие отбора большинство возникших мутаций быстро утрачивается, однако некоторые из них могут сохраниться в ряде поколений. Исчезновению мутантных генов из популяции противостоит действие мутационного процесса, в результате которого образуются повторные мутации.
Генетическая структура популяций формируется и изменяется под действием естественного и искусственного отбора. Действие естественного отбора состоит в том, что преимущественное размножение имеют особи с высокой жизнеспособностью, скороспелостью, плодовитостью и т. д., т. е. более приспособленные к условиям окружающей среды. При искусственном отборе определяющее значение имеют признаки продуктивности.
В.И. Власов отмечает, что естественный отбор идет на всех этапах онтогенеза популяции - от образования гамет до взрослого организма. При этом он существенно влияет на темпы искусственного отбора вследствие противоположного действия при селекции на высокий уровень развития продуктивных признаков, несвойственный видовым биологическим границам. Исходя из этого, при отборе животных необходимо учитывать не только продуктивные признаки, но и признаки приспособленности к условиям окружающей среды.
По С.М. Гершензону, критерием интенсивности естественного отбора служит разность приспособленности сравниваемых групп, называемая коэффициентом отбора и выражаемая в долях единицы. Например, если вероятность оставления потомства особями с генотипом аа на 10% меньше, чем особями с генотипом АА или Аа, то приспособленность этих групп для особей АА и Аа равна 1, для особей аа - 0,9.
С точки зрения ветеринарной генетики имеет значение эффективность отбора против вредных мутаций, прежде всего рецессивного типа. Анализ показывает, что высокие частоты рецессивного мутантного гена путем отбора могут быть быстро снижены до низких значений. Чтобы снизить частоту летального гена, например с 0,3 до 0,2, достаточно двух поколений.
Частота гомозигот (аа) по мутантному гену зависит от частоты гетерозиготных животных в популяции. Выявление этих гетерозигот и элиминация их соответственно будут снижать частоту генетических аномалий, обусловленных мутантным геном, что особенно важно при высокой частоте мутаций.
Генетическая структура популяции может изменяться в силу случайных генетико-автоматических процессов (по Н.П. Дубинину) или дрейфа генов (по С. Райту). Наблюдения показывают, что наиболее интенсивно дрейф генов протекает в малых популяциях. Например, известны случаи высокой концентрации редких мутаций в малочисленных изолированных популяциях крупного рогатого скота и других видов животных, связанные, очевидно, с генетико-автоматическими процессами. Распространение мутаций в разных популяциях животных может произойти в результате миграций.
Спаривание животных, находящихся в родственных отношениях, называют инбридингом. Родственное спаривание, или инбридинг, - метод подбора, используемый в племенном животноводстве для закрепления ценных наследственных признаков того или иного животного в последующих поколениях. У родственных между собой животных наблюдается сходство по определенным парам аллелей, которые они получили от общего предка. Это сходство тем больше, чем ближе степень родства.
Каждое животное в генотипе имеет аллельные гены, как в гомозиготном, так и в гетерозиготном состоянии. В гетерозиготе обычно находятся вредные мутантные рецессивные гены. При инбридинге возрастает вероятность слияния тождественных гамет, несущих мутантные гены в гетерозиготном состоянии, и перехода их в гомозиготное состояние. Эта вероятность пропорциональна степени родства спариваемых животных.
Таким образом, в результате применения инбридинга происходит изменение генных частот, возрастает вероятность выщепления рецессивных гомозигот, что является причиной инбредной депрессии, выражающейся в снижении жизнеспособности, плодовитости животных, рождении аномальных особей.
Инбридинг, как правило, был комплексным - одновременно на двух (1-я цифра) или на трех (2-я цифра) предков.
Имбредная депрессия по показателям, характеризующим продуктивность и жизнеспособность животных, не является фатальным спутником родственного спаривания.
Имеется множество примеров того, когда при инбридинге разных степеней, в том числе и близких, отрицательных последствий не наблюдали.
Н.П. Дубинин в этой связи отмечает, что «линия ухудшается, пока в ней идут процессы последовательного накопления вредных рецессивных генов, переходящих в гомозиготное состояние. Когда же наступает более или менее выраженное завершение этого процесса, линии по своим свойствам становятся относительно константными и могут в таком устойчивом состоянии сохраняться длительно. Изменить генотип таких линий могут лишь новые накапливающиеся в них мутации». Однако, подчеркивает академик, «многие линии при инбридинге, конечно, гибнут, ибо в них в гомозиготное состояние переходят летальные и полулетальные гены». Поэтому инбридинг применяется как метод индивидуальной селекции для перевода в гомозиготное состояние ценных генов выдающихся животных.
В ходе длительной эволюции животных наряду с полезными мутациями, подхватываемыми отбором, в популяциях или породах накопился определенный спектр генных и хромосомных мутаций. Каждое поколение популяции наследует этот груз мутаций, и в каждом из них возникают новые мутации, часть которых передается последующим поколениям.
Очевидно, что большая часть вредных мутаций отметается естественным отбором или элиминируется в процессе селекции. Это прежде всего доминантные генные мутации, фенотипически проявляющиеся в гетерозиготном состоянии, и количественные изменения наборов хромосом. Рецессивно действующие генные мутации в гетерозиготном состоянии и структурные перестройки хромосом, заметно не влияющие на жизнеспособность их носителей, могут проходить сквозь сито селекции. Они формируют генетический груз популяции. Таким образом, под генетическим грузом популяции понимают совокупность вредных генных и хромосомных мутаций. Различают мутационный и сегрегационный генетический груз. Первый формируется вследствие новых мутаций, второй - в результате расщепления и перекомбинирования аллелей при скрещивании гетерозиготных носителей «старых» мутаций.
Частота летальных, полулетальных и субвитальных мутантных генов, передающихся из поколения в поколение в форме мутационного генетического груза, из-за трудности идентификации носителей не поддается точному учету. Мортон и Кроу предложили форму расчета уровня генетического груза в количестве летальных эквивалентов. Один летальный эквивалент соответствует одному летальному гену, обусловливающему смертность с 10%-ной вероятностью, двум летальным генам при 50%-ной вероятности смерти и т. д. Величина генетического груза по формуле Мортона:
log eS = A + BF,
где S- часть потомства, оставшаяся в живых;
А - смертность, измеряемая летальным эквивалентом в популяции при условии случайных спариваний (F= 0), плюс смертность, обусловленная внешними факторами;
В - ожидаемое увеличение смертности, когда популяция становится полностью гомозиготной (F= 1);
F - коэффициент инбридинга.
Уровень генетического груза можно определять на основании фенотипического проявления мутаций (уродства, врожденные аномалии обмена и т. д.), анализа типа их наследования, частоты в популяции.
Н.П. Дубинин предлагает определять генетический груз популяции путем сравнения частот мертворожденных в родственных и неродственных подборах родительских пар. При этом следует иметь в виду, что при высокой частоте гетерозигот по рецессивным летальным и полулетальным мутантным генам рождение животных с аномалиями необязательно должно быть связано с инбридингом близких и умеренных степеней. Общий предок (источник мутации) может находиться и в отдаленных рядах родословной. К примеру, бык Трувор 2918 - гетерозиготный носитель мутантного рецессивного гена, находился в V, VI, VII рядах предков в совхозе «Красная Балтика», но при использовании его праправнука Автомата 1597 на родственных ему коровах наблюдались массовые случаи рождения бесшерстных телят.
Эти данные в определенной мере характеризуют уровни генетического груза по отдельным мутантным генам в конкретных популяциях крупного рогатого скота.
Хромосомные мутации являются составной частью генетического груза. Учет их ведется прямым цитологическим методом. По результатам многочисленных исследований основной компонентой груза аберраций хромосом у крупного рогатого скота являются робертсоновские транслокации, а у свиней - реципрокные. Наиболее распространенной мутацией у крупного рогатого скота оказалась транслокация 1/29 хромосомы. Размах изменчивости частоты этой аберрации, по нашим данным, в популяциях палево-пестрого скота составлял от 5 до 26%.
Таким образом, концепция генетического груза в свете современных достижений цитогенетики должна быть расширена. Сейчас, когда известен широкий спектр аберраций хромосом и установлено строгое наследование отдельных из них (транслокации и инверсии), представляется целесообразным учитывать их наряду с вредными мутациями генов как составляющую часть генетического груза.
Существование в популяциях наследственной изменчивости, прежде всего мутаций в гетерозиготном состоянии, позволяет им быстро приспосабливаться к новым условиям среды за счет изменения генетической структуры. Мутационный процесс ведет также к образованию в популяциях генетического полиморфизма - разнообразия частот аллелей, гомозигот по доминантным, гетерозигот или гомозигот по рецессивным генам. Полиморфизм является механизмом, поддерживающим существование популяций. Если, например, гетерозиготность обеспечивает лучшую приспособляемость к изменившимся условиям среды, то идет отбор в пользу гетерозигот, что приводит к сбалансированному полиморфизму - воспроизведению в популяции из поколения в поколение определенного соотношения различных генотипов и фенотипов. Процессы, обеспечивающие способность популяции сохранять свою генетическую структуру, называют генетическим гомеостазом.
В генетике выделяют два класса признаков - качественные и количественные. Они различаются по характеру изменчивости и особенности наследования. Качественные признаки характеризуются прерывистой, а количественные - непрерывной изменчивостью. Первые из них дают четкие границы при расщеплении на доминантные или рецессивные признаки. Это связано с тем, что каждый из них обычно контролируется одним аллельным геном. Количественные признаки не дают четких границ расщепления при разных вариантах скрещивания, хотя отличаются от качественных более высокой степенью изменчивости. Особенностью количественных признаков является сложный характер наследования. Каждый из них детерминируется не одним, а множеством локусов в хромосомах. Такой тип наследования, когда один признак обусловливается многими генами, носит название полигенного. Уровень развития количественного признака зависит от соотношения доминантных и рецессивных генов, других генетических факторов и степени модифицирующего действия факторов внешней среды. Изменчивость по количественному признаку в популяции складывается из генетической и паралогической (внешнесредовой) изменчивости.
Понятие о наследуемости признаков и коэффициенте наследуемости. Разные количественные признаки имеют неодинаковую степень генетической изменчивости, и условия внешней среды оказывают различное воздействие на уровень фенотипического проявления того или иного признака. При отборе животных важнейшее значение имеет знание того, в какой степени будет совпадение уровней развития количественного признака у родителей и потомства или в какой степени потомки унаследуют количественные хозяйственно полезные признаки или патологические признаки родителей.
К хозяйственно полезным признакам относят молочность, содержание жира и белка в молоке коров, настриг шерсти у овец, яйценоскость у кур, прирост живой массы, плодовитость и др. Повышение уровня развития хозяйственно полезных признаков достигается постоянным отбором лучших индивидуумов для воспроизводства. Эффективность отбора по этим признакам зависит от степени их наследуемости, взаимосвязи между ними, разности между средним значением признака отобранной группы и средним по стаду (селекционный дифференциал) и интервала между поколениями.
Величины коэффициентов наследуемости зависят от природы признака. Так, среднее значение А2 по удою равно 0,25, жирномолочности - 0,38, живой массе у овец - 0,35, выходу чистой шерсти - 0,55, плодовитости у крупного рогатого скота - 0,08 и т. д.
Селекционной практикой и специальными исследованиями накоплены данные, свидетельствующие о том, что в некоторых случаях уровень развития одного или нескольких признаков у потомства превосходит степень выраженности этих признаков у лучшего из родителей. Такое явление, названное гетерозисом, не вписывается в обычные рамки наследования признаков. Для его объяснения предложены разные гипотезы:
1) гетерозиготного состояния по многим генам;
2) взаимодействия доминантных благоприятных генов;
3) сверхдоминирования, когда гетерозиготы превосходят гомозиготы.
Н.В. Турбин предложил теорию генетического баланса, в основе которой лежит сложный характер причинно-следственных связей между наследственными факторами и признаками.
Н.Г. Дмитриев и И.JI. Гальперин отмечают, что главную причину возникновения гетерозиса надо искать в особенностях эволюции вида, породы, линии. При этом следует иметь в виду, что все в природе направлено на сохранение жизни. Проявление гетерозиса зависит от генетической природы признака. Так, при межпородном или межлинейном скрещивании гетерозис в большей степени проявляется в отношении признаков, имеющих Низкую степень наследуемости
Что касается признаков, имеющих среднюю или высокую наследуемость, то гетерозис по ним Чаще всего проявляется слабо, и гибриды занимают обычно промежуточное положение.
При наличии истинного гетерозиса величина индекса или больше 100%. Если же величина гетерозиса меньше 100% или имеет знак «минус», то правильнее говорить о лучшей или худшей комбинационной способности линий. Скрещивание последних по определенной схеме обеспечивает в гибридном потомстве лучшее развитие одного признака от отца, а другого от матери, хотя этот признак у гибрида по своему развитию не превосходит лучшую родительскую форму.




По вертикали:
Так называется популяция растений искусственно созданная человеком?
Как называется метод при котором проводят различные скрещивания организмов?
По горизонтали:
В основе этого метода, который используется до сих пор лежит концепция разработанная еще Ч.Дарвиным.
Так называется популяция животных искусственно созданная человеком?
Так называется популяция микроорганизмов искусственно созданная человеком?








Одомашнивание началось более 10 тыс. лет назад. Его центры в основном совпадают с центрами многообразия и происхождения культурных растений. Одомашнивание способствовало резкому повышению уровня изменчивости у животных. Гибридизация и индивидуальный отбор являются основными методами в селекции животных. Массовый отбор практически не применяется из-за небольшого количества особей в потомстве. В селекции животных применяют 2 вида гибридизации.


Инбридинг – родственная гибридизация. Скрещивание между братьями и сестрами или между родителями и потомство ведет к гомозиготности и часто сопровождается ослаблением животных, уменьшению их устойчивости к неблагоприятным условиям, снижению плодовитости. Тем не менее инбридинг применяют с целью закрепления в породе характерных хозяйственно ценных признаков.

Аутбридинг – неродственная гибридизация. Это скрещивание сопровождается строгим отбором, что позволяет усиливать и поддерживать ценные качества породы. Сочетание – родственной и неродственной гибридизации широко применяется селекционерами для выведения новых пород животных. Важным направлением в селекции животных является направление гетерозиса. Особенно широко явление гетерозиса применяется например в птицеводстве, например при получении бройлерных цыплят.

Собаки и волки скрещиваются довольно свободно. Волк – это пугливое животное с особенным поведение и развитым охотничьим инстинктом. Челюсти у него гораздо мощнее, чем у собаки. Поведение гибридов волка и собаки непредсказуемо. Для того, чтобы приручить животное, обязательно нужна дрессировка
Сегодня мы попробуем провести «виртуальную» экскурсию среди некотрых пород домашних животных, а заодно и вспомним основные методы селекции животных.
1. Аутбридингом получены например, собаковолк

2. Инбридингом получены:
Тигролев – это помесь самца тигра и самки льва. Они имеют склонность к карликовости и обычно по размерам меньше своих родителей. Самцы бесплодны, в то время как самки порой могут приносить потомство.

Лигр – это помесь самца льва и самки тигра. Они являются самыми крупными из семейства кошачьих в мире. Самцы бесплодны, в то время как самки порой могут приносить потомство.

Левопард – это результат скрещивание самца леопарда с самкой льва. Голова животного похожа на голову льва, в то время как остальное тело больше напоминает леопарда. По размерам левопарды крупнее обычных леопардов, они любят карабкаться по деревьям и плескаться в воде.

3. У домашних животных наблюдается явление гетерозиса: при межпородных или межвидовых скрещиваниях у гибридов первого поколения происходит особенно мощное развитие и повышение жизнеспособности. Классическим примером проявления гетерозиса является мул – гибрид кобылы и осла. Это сильное, выносливое животное, которое может использоваться в значительно более трудных условиях, чем родительские формы.

Тестовые задания по теме « Селекция»
1. Аутбридинг - это:
1)скрещивание между неродственными особями одного вида;
2)скрещивание различных видов;
3)близкородственное скрещивание;
4)нет верного ответа.
2. Гибриды, возникающие при скрещивании различных видов:
1)отличаются бесплодностью;
2)отличаются повышенной плодовитостью;
3)дают плодовитое потомство при скрещивании с себе подобными;
4)всегда бывают женского пола.
3. Центром происхождения культурных растений считаются районы, где:
1)обнаружено наибольшее число сортов данного вида;
2)обнаружена наибольшая плотность произрастания данного вида;
3)данный вид впервые выращен человеком;
4)нет верного ответа.
4. Многообразие пород кошек является результатом:
1)естественного отбора 2)искусственного отбора
3)мутационного процесса 4)модификационной изменчивости
5. Особенностью селекции животных является:
1)неприменимость методов гибридизации;
2) неспосбность к бесполому размножению
3) отсутствие мутаций
6. основным критерием для установления родства между видами является:
1) внешнее сходство
2)генетическое сходство
3) общие центры происхождения
7. Одомашнивание является начальным этапом:а) селекции растений, животных; б) селекции растений; в) гибридизации; г) селекции животных.

А. Сорт. Б. Породы. В. Штамм. Г. Раса.
8. Назвать фактор, который является главным в создании новых сортов и пород..
А. Наследственность. Б. Изменчивость. В. Искусственный отбор. Г. Естественный отбор.
9. Назвать соотношение полов во время полового размножения.
А. 2: 2. Б. 1: 1. В. 3: 1. Г. 4: 1.
10. Назвать стадию онтогенеза, которой заканчивается формирование нервной трубки у млекопитающих.
А. Бластула. Б. Гаструла. В. нейрулы. Г. Зародыш.
11. Отметьте особенность, не характерная для мутации.
А. Возникают внезапно, резко. Б. Имеют адаптивное значение.
В. Наследуются. Г. Не имеют массовый характер.
12. Назовите половые хромосомы курицы.
А. хх. Б. Ху. В. хо. Г. Отсутствуют.
в качестве самостоятельной науки экология начала развиваться 5. направление движения естественному отбору диктует 6. Факторы окружающей среды, воздействует на организм 7. Группа экологических факторов, обусловленная влиянием живых организмов 8. Группа экологических факторов, обусловлена влиянием живых организмов 9 . Группа экологических факторов, обусловленная влиянием неживой природы 10. Фактор неживой природы, дающий толчок сезонным изменениям в жизни растений и животных. 11. способность живых организмов иметь свои биологические ритмы в зависимости от длины светового дня 12. Самый значимый для выживания фактор 13. Свет, химический состав воздуха, воды и почвы, атмосферное давление и температура относиться к факторам 14. строительство железных дорог, распашка земель, создание шахт относяться 15. Хищничество или симбиоз относиться к факторам 16. растения длинногодн обитают 17. растения короткого дня обитания 18.растени тундры относиться 19.РАстения полупустынь,степей и пустынь относиться 20. Характерный показатель популяции. 21. Совокупность всех видов живых организмов, населяющих определенную территорию и взаимодействующих между собой 22. Наиболее богатая видовым разнообразием экосистема нашей планеты 23. экологическая группа живых организмов, создающих органические вещества 24. экологическая группа живых организмов,потребляющие готовые органические вещества, но не проводящих минерализации 25. экологическая группа живых организмов,потребляющих готовые органические вещества и способствующих полному превращению их в минеральные вещества 26 . полезной энергии на следующий трофический(пищевой) уровень переходит 27 . консументы I порядка 28. консументы IIили III порядка 29. мера чувствительности сообществ живых организмов к изменениям определенных условий 30.способность сообществ (экосистем или биогеоценозов) поддерживать свое постоянство и противостоять извенению условий окружающей среды 31. низкая способность к саморегуляции, видовое разнообразие, использование дополнительных источников энергии и высокая продуктивность характерны для 32. искусственный биоценоз с наибольшей интенсивностью обмена веществ на единицу площади. с вовлечением круговорот новых материалов и выделением большого количества неутилизируемых отходов характерны для 33. пахотными землями занято 34. города занимают 35. оболочка планеты, заселенная живыми организмами 36. автор учении о биосфере 37. верхняя граница биосферы 38. граница биосферы в глубинах океана. 39 нижняя граница биосферы в литосфере. 40 . международная неправительственная организации, созданная в 1971 году, совершающая наиболее действенные акции в защиту природы.
А 1. Критерий вида, учитывающий совокупность факторов среды, в которой существует вид:Морфологический;
Физиологический;
Географический;
Экологический.
А 2. Элементарная структура, на уровне которой проявляется действие естественного отбора – это:
Отдельный организм;
Популяция;
Биоценоз;
А 3. Естественный отбор в отличие от искусственного:
Способствует сохранению полезных для организма признаков;
Обеспечивает сохранение особей с полезными для человека признаками;
Направлен на создание и улучшение сортов и пород;
Действует с момента появления земледелия и скотоводства.
А 4. К результатам эволюции относят:
Наследственную изменчивость;
Борьбу за существование;
Приспособленность;
Естественный отбор.
А 5. Примером палеонтологических доказательств эволюции служат:
Отпечатки и окаменелости древних форм жизни;
Наличие у китов рудиментов конечностей;
Признаки пресмыкающихся в строении утконоса;
Признаки сходства у зародышей млекопитающих и рыб на ранних стадиях развития.
А 6. Примером ароморфоза у хвойных растений служит возникновение у них:
А 7. К дегенерации относят:
Утрату большинства органов корнеголовым раком саккулиной;
Появление четырёхкамерного сердца у птиц;
Появление шерстного покрова у млекопитающих;
Формирование плоской формы тела у скатов.
А 8. У человека в отличие от человекообразных обезьян:
Имеются мимические мышцы;
Имеются передние конечности с ногтями;
Постоянная температура тела;
Позвоночник имеет четыре изгиба.
А 9. Примером биологического фактора эволюции человека может служить:
Способность к трудовой деятельности;
Использование одежды;
Общение с помощью устной и письменной речи;
Способность передавать приобретённые признаки по наследству.
А 10. Выведением новых сортов и пород растений и животных занимается…
Селекция
Генетика
Физиология
Цитология
помогите пожалуйста??
1. Выведением новых сортов растений и пород животных занимается:А – генетика;
Б – селекция;
В – агробиология;
Г – ботаника.
2. Наследственность – это свойство организмов:
А – взаимодействовать со средой обитания;
Б – реагировать на изменения окружающей среды;
В – передавать свои признаки и особенности развития потомству;
Г – приобретать новые признаки в процессе индивидуального развития.
3. Для изучения характера наследования нескольких признаков рядом поколений растений и животных проводят скрещивание:
А – моногибридное;
Б – анализирующее;
В – полигибридное;
Г – близкородственное.
4. «Расщепление по каждой паре признаков идет независимо от других пар признаков» – это формулировка:
А – первого закона Менделя;
Б – закона Моргана;
Г – второго закона Менделя;
Д – третьего закона Менделя.
5. Появление в первом гибридном поколении особей с одинаковым генотипом является проявлением:
А – закона расщепления;
Б – закона независимого наследования;
В – правила единообразия;
Г – закона сцепленного наследования.
6. На рис. изображены родительские формы, у которых тюльпанов красный цвет лепестков доминирует над белым. Каким будет генотип потомства по этому признаку, если родительский организм с доминантными признаками гомозиготен?
А – АА;
Б – аа;
В – Ааа;
Г – Аа.
7. По рис. определите генотип потомства (F1) морских свинок, если известно, что родительская особь с черной и короткой шерстью гетерозиготна по обоим признакам:
А – АаВв;
Б – аавв;
В – ааВв;
Г – Аавв.
8. Генотип – это совокупность:
А – внешних признаков организма;
Б – внутренних признаков организма;
В – генов, полученных потомством от родителей;
Г – реакций организма на воздействие среды.
9. Промежуточный характер наследования признаков проявляется в том случае, когда:
А – наблюдается изменение условий среды обитания;
Б – происходят сезонные изменения в природе;
В – гетерозиготные особи внешне не отличаются от гомозиготных;
Г – гетерозиготные особи внешне отличаются от гомозиготных.
10. Гены, расположенные в одной хромосоме:
А – наследуются независимо;
Б – попадают в разные половые клетки в процессе мейоза;
В – наследуются вместе;
Г – дают расщепление в потомстве в соотношении 3:1.
11. Какой буквой обозначен фенотип организма, изображенного на рис.
Б – AaBbCc;
В – AbC;
Г –
12. Скрещивание особей, различающихся по двум парам признаков, называют:
А – полигибридным;
Б – анализирующим;
В – дигибридным;
Г – моногибридным.
13. С генетической точки зрения наследственные заболевания у человека представляют собой:
А – модификационные изменения;
Б – изменение фенотипа, не связанное с изменением генотипа;
В – мутации;
Г – реакцию на изменения среды обитания, не зависящую от генотипа.
14. В основе цитогенетического метода изучения наследственности человека лежит исследование:
А – родословной семьи;
Б – распространение признака в большой популяции людей;
В – хромосомного набора, отдельных хромосом;
Г – развития признаков у близнецов.
15. Изменение последовательности расположения нуклеотидов в молекуле ДНК называют:
А – генными мутациями;
Б – хромосомными мутациями;
В – соматическими мутациями;
Г – комбинативной изменчивостью.
16. Границы, в пределах которых возможны модификации того или иного признака, называют:
А – приспособленностью;
Б – нормой реакции;
В – изменчивостью;
Г – раздражимостью.
17. Под воздействием генотипа и условий среды обитания формируется:
А – норма реакции;
Б – наследственность;
В – фенотип;
Г – приспособленность.
18. Выделение из исходного материала целой группы особей с необходимыми для селекционера признаками называют:
А – естественным отбором;
Б – массовым отбором;
В – индивидуальной формой искусственного отбора;
Г – стихийным отбором.
Определение понятия «вид» представляется очень трудным делом.
Мы будем рассматривать вид как исторически сложившуюся совокупность организмов, занимающих определенный ареал обитания, и характеризующихся общностью происхождения, сходной системой приспособлений к уровням среды и воспроизведением в поколениях основных адаптивных черт и признаков.
Организмы одного вида обладают характерными для данного вида фенотипом и генотипом, отличными от таковых у организмов другого вида.
Вид занимает определенный ареал. Ареал распространения одних видов оказывается широким, и такие виды являются обычно политипическими - включают несколько географических рас, или подвидов. Другие виды имеют значительно более ограниченный ареал, они, как правило, не образуют географических рас и являются монотипическими. Особи, составляющие вид, не образуют собой неизменную однородную массу. Каждый организм вида, обладая общими и характерными чертами, имеет и свои индивидуальные генотипические особенности, представляющие в совокупности наследственную изменчивость вида, или, как иногда называют, «генофонд» вида.
Вид, представляя единую совокупность организмов, расчленяется на отдельные популяции. Популяцией называют совокупность свободно скрещивающихся особей одного вида, характеризующихся общностью местообитания и приспособления к данным условиям существования. Популяция складывается под влиянием условии существования на основе взаимодействия факторов наследственности, изменчивости, и отбора. Образование популяций является своеобразным способом «пригонки» вида к конкретным условиям его существования. Породы животных и сорта растений, создаваемые искусственным отбором, также представлены отдельными популяциями.
Процессы формирования популяций и их динамики составляют микроэволюцию. Возникновение новых видов начинается с дивергенции - расчленения вида на отдельные, нескрещивающиеся или изолированные группы организмов. Популяция является своего рода «кузницей», в которой естественный отбор творит новые формы.
В природе популяции каждого вида характеризуются генетическим разнообразием. Но мы часто этого не замечаем. Особи популяции и вида внешне кажутся нам относительно однообразными. Это относительное однообразие животных и растений, позволяющее систематикам относить животных и растения к определенным видам, подвидам, расам, создается естественным отбором. Отбор обеспечивает не только разнообразие, но и однообразие внутри вида.
Однако указанное однообразие относится лишь к основным типичным чертам, признакам и свойствам организмов данной популяции. Как только мы начнем детально генетически анализировать состав популяции, разлагая ее на отдельные линии, так сейчас же обнаружим огромную генотипическую изменчивость. При этом оказывается, что каждой популяции, в пределах которой особи длительное время скрещивались друг с другом, на ограниченной территории при данных климатических условиях присущ свой характер изменчивости.
Источниками наследственной изменчивости в популяции являются мутационная и комбинативная изменчивости. Действие генетических законов в популяции представляет предмет исследования генетики популяций.
В природе не существует двух организмов, абсолютно сходных между собой по генотипу. Зоологи или ботаники, изучающие биологические процессы у какой-либо группы особей, всегда имеют дело с наследственно разнородной группой организмов. Но поскольку ими изучается фенотип организмов, они вправе пренебрегать генетическим разнообразием своего материала.
Изучение популяции можно вести чисто описательным методом. В этом случае определяют фенотипическую характеристику форм популяции, ее биологические особенности, выясняют условия существования и взаимоотношение организмов, цепи питания, конкуренцию, динамику численности по годам и зависимость ее от разных факторов. Популяции обособляются и складываются в результате действия многих факторов: способа размножения, характера изменчивости, изменения численности особей, темпа и направления отбора, климатической, географической и физиологической изоляции. Основным из них является отбор признаков, обеспечивающих процесс воспроизведения поколений, т. е. размножение. Очевидно, что при разных способах размножения возникновение и поддержание популяций осуществляется разными путями, в чем можно убедиться, сравнивая популяции перекрестно оплодотворяющихся (аллогамных) и самооплодотворяющихся (автогамных) организмов.
Для существования популяции имеют первостепенное значение различные типы наследственной изменчивости: генные мутации, хромосомные перестройки и полиплоидия. Ненаследственные изменения могут играть лишь ограниченную роль. Организмы, генотипически различающиеся, например, по одному гену, морфологически могут не отличаться друг от друга, но обладать различными физиологическими особенностями (жизнеспособностью, длительностью развития, плодовитостью). Генетические методы дают возможность составить более полное представление о наследственных потенциях популяции, ее приспособительных особенностях и направлении эволюции.
Зачинателями изучения генетической надо признать селекционеров, ибо для того, чтобы выявить разнообразие генотипов в популяциях, необходимо было изолировать, подобрать для скрещивания отдельные пары родителей и затем исследовать их потомство в ряду поколений. Именно так поступали селекционеры, создававшие различные сорта и породы. Однако научные основы генетического изучения популяции могли быть заложены только после открытия Г. Менделя, установившего количественные закономерности наследования.
Первое научное изучение популяции, совместившее в себе генетические и статистические методы, было предпринято датским физиологом растений и генетиком В. Иоганнсеном. Его классическая работа «О наследовании в популяциях и чистых линиях», опубликованная в 1903 г., положила начало генетическому изучению популяций. Как часто бывает в науке, классическое открытие делается на элементарном, на первый взгляд, явлении и при использовании простой методики. Так и в данном случае В. Иоганнсен избрал объектом исследования популяции не перекрестноопыляющиеся, а самоопыляющиеся растения - ячмень, фасоль и горох. Методически это упрощало работу, поскольку каждую такую популяцию можно было легко разложить на группы потомков отдельных особей, т. е. произвести выделение отдельных, «чистых», линий. «Чистой линией, - писал он, - я называю индивиды, которые происходят от одного самоопыляющегося индивида. Отсюда ясно, что популяция абсолютных самоопылителей состоит только из чистых линий, индивиды которых в природе могут быть смешаны, но не могут быть затронуты скрещиванием».
В качестве признаков были взяты вес и размеры семян. Эти количественные признаки определяются действием многих генов, т. е. определяются полигенно, и в сильной степени подвержены изменчивости под влиянием факторов внешней среды - состава почвы, климата, способа посадки растений и т. п. Поэтому для установления характера их наследования необходимо использование математических методов анализа изменчивости.
По этим признакам имеется ярко выраженная модификационная, или паратипическая, изменчивость. О значении этой изменчивости для эволюции в биологии имелись различные точки зрения. Сторонники теории наследования приобретенных свойств считали, что изменения, вызванные воздействием факторов внешней среды, наследуются, передаются потомству. Противники этой теории отрицали наследование модификационных изменений. Решение этого спора в пользу последних имело принципиальное значение, так как отбор организмов по фенотипу без выявления наследственных потенций имел до этого широкое распространение в селекции и тормозил выведение пород животных и сортов растений.
Иоганнсен провел взвешивание семян одно сорта фасоли и построил вариационный ряд по этому показателю. Вес семян оказался изменчивым в пределах от 150 до 750 мг. В дальнейшем семена весом от 250 до 350 мг были высеяны отдельно от семян весом 550- 650 мг. С каждого выросшего растения вновь были взвешены семена. Так как фасоль является самоопыляющимся растением, то генотип семян с одного растения должен быть одинаковым, а у семян от разных растений могут быть генотипические различия. Поэтому тяжелые семена (550-650 мг) и легкие (250-350 мг), выбранные из сорта, представляющего популяцию, дали растения, семена которых значительно различались по весу. Средний вес семян на растениях, выросших из тяжелых семян, составил 518,7 мг, а на растениях, выросших из легких семян, - 443,4 мг. Этим было показано, что сорт-популяция фасоли состоит из генетически различных растений, каждое из которых может стать родоначальником чистой линии.
На протяжении 6-7 поколений Иоганнсен отбирал также тяжелые и легкие семена с каждого растения в отдельности, т. е. производил отбор в пределах чистых линий. При таком отборе ни в одной линии не произошло сдвига в ряду поколений в сторону тяжелых или легких семян. Следовательно, изменчивость по весу семян внутри чистой линии была ненаследственной, модификационной.
В результате своих исследований Иоганнсен пришел к следующим выводам: 1) «отбор в популяции вызывает… больший или меньший сдвиг - в направлении отбора - того среднего признака, вокруг которого, флуктуируя, варьируют соответствующие индивиды» и 2) «внутри чистых линий регрессия (степень сходства признака потомства с материнским) была… полной; отбор внутри чистых линий не вызывал никакого смещения типа».
Как мы видим, популяция автогамных растений состоит из генотипически разнородных линий. Растения такой популяции не скрещиваются между собой и не обмениваются наследственной информацией. В этом случае существование популяции основывается на строгом естественном отборе линий определенного генотипа, на общности приспособительных механизмов к однотипным условиям внешней среды. Иначе говоря, изменение популяции автогамных растений и животных осуществляется отбором определенных наследственно различающихся линий и клонов, имеющих адаптивные преимущества.
При самооплодотворении отдельный организм может быть зачинателем новой расы, подвида и вида, а также сорта или породы. Например, новый сорт пшеницы может быть выведен от одного зерна, отобранного из популяции.
Однако, говоря о высокой гомозиготности в чистых линиях, следует иметь в виду, что даже чистые линии не могут быть абсолютно гомозиготными по следующим обстоятельствам. Во-первых, облигатных (абсолютных) самоопылителей-растений не существует. В популяциях самоопылителей, например пшениц, томатов и др., всегда с той или иной частотой встречаются растения с открытым цветением и перекрестным опылением. В силу этого между чистыми линиями в популяциях хотя и редко, но совершаются процессы скрещивания и соответственно обмена наследственной информацией. Во-вторых, у растений-самоопылителей встречаются мутации, препятствующие самоопылению (несовместимость). В-третьих, в чистых линиях самоопылителей даже за одно поколение возникает весьма ощутимое количество разнообразных мутаций, которые нарушают гомогенность чистой линии.
Вследствие указанных причин сорта растений-самоопылителей при воспроизведении в производстве могут утрачивать часть своих сортовых качеств и требуют постоянного контроля, на чем и основана необходимость сортообновления.
При вегетативном размножении агамных организмов, не имеющих полового процесса или вторично его утративших (некоторые простейшие, грибы, водоросли и др.), объектом отбора в популяции являются отдельные клоны. Генетическая целостность (интеграция) таких клонов в популяции является очень низкой в силу невозможности осуществления скрещиваний между особями разных клонов, но такие популяции, по-видимому, все же существуют в природе и поддерживаются отбором на основе симбиотических отношений разных генотипов.
У перекрестно оплодотворяющихся организмов в природе популяция формируется на основе свободного скрещивания разнополых особей с разными генотипами, т. е. на основе панмиксии. При этом наследственная структура следующего поколения воспроизводится на основе разнообразных сочетаний различных гамет при оплодотворении. Отсюда вытекает, что численность особей того или иного генотипа в каждом поколении будет определяться частотой встречаемости разных гамет, произведенных генотипически различными родительскими организмами. Это значит, что признаки и свойства сохраняются и распределяются в популяции на основе закономерностей изменения частоты распространения генов. В основе таких изменений лежат закономерности наследования, открытые Г. Менделем и Т. Морганом. Знание этого позволило вывести правила распространения генов в панмиктической популяции.
Очевидно, те организмы, генотипы которых наилучшим образом обеспечивают приспособление к условиям существования, произведут большее число соответствующих гамет, чем менее приспособленные. Следовательно, частота (встречаемость) того или иного гена в популяции также будет определяться естественным отбором.
Сообщество свободноскрещивающихся генотипически различных организмов внутри вида некоторые генетики называют менделевской популяцией. Мы предпочитаем называть ее панмиктической, поскольку ее существование определяется не только законами Менделя, но и взаимодействием всех факторов эволюции, обеспечивающих свободу скрещивания организмов внутри популяции. Разнообразие генотипов панмиктической популяции - это результат мутационной и комбинативной изменчивости. Вновь возникшая мутация, чтобы стать достоянием популяции, должна сохраниться и размножиться, т. е. оказаться в составе генотипов целого ряда организмов. Любая мутация в популяции имеет свою судьбу.
Вследствие распространения большого количества разнообразных мутаций в популяции генотипы организмов оказываются насыщенными различными мутациями, которые чаще всего находятся в гетерозиготном состоянии. Так, например, что число растений, гетерозиготных по некоторым мутациям, может составлять в популяции довольно высокий процент. По мере повышения концентрации мутаций в популяции они переходят в гомозиготное состояние.

Огромная насыщенность популяции мутантными генами характерна не только для культурных растений и домашних животных, но, как впервые показал С. С. Четвериков, и для природных популяций. При этом в популяции встречаются мутации, различающиеся как по своей генетической природе (генные мутации и хромосомные перестройки), так и по своему фенотипическому проявлению.
Чтобы иллюстрировать расчленение панмиктической популяции под действием отбора, рассмотрим модельный опыт с искусственно созданной гибридной популяцией, поставленный американскими генетиками Д. Джонсом и Е. Истом. Эти два исследователя скрещивали две разновидности табака, различавшиеся по длине венчика (короткий и длинный). Растения первого поколения были скрещены между собой, а из второго поколения были взяты две линии А и В со сходной изменчивостью по этому признаку.
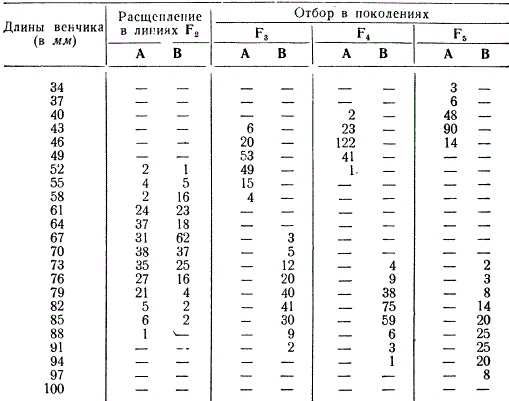
Длина венчика определяется многими генами, и поэтому в F 2 она колебалась в этих линиях в пределах от 52 до 88 мм, В дальнейшем в потомстве взятых линий был произведен отбор в течение трех поколений: в линии А - на короткий венчик, а в линии В - на длинный венчик. В каждом поколении внутри обеих линий скрещивали отобранные формы: в линии А - с коротким венчиком, а в линии В - с длинным венчиком. Как мы видим, уже в пятом поколении линии А и В настолько сильно различались, что между ними не было никакого захождения (трансгрессии), т. е. максимальная длина венчика в линии А была меньше минимальной длины в линии В.
Следовательно, путем отбора и скрещивания отбираемых форм удается создать линии с иным выражением признака, чем у исходной популяции: отбор расчленяет популяцию на различные генотипы. В данном опыте искусственный отбор был проведен по одному признаку с преднамеренным скрещиванием растений. В природе же естественный отбор осуществляется по многим признакам и либо сохраняет и поддерживает популяцию в целостном состоянии, либо разлагает ее соответственно конкретным условиям существования.
Изучение генетики популяций ведется разными методами, основными из которых являются – цитогенетический, эколого-физиологический, математический.
Первые два метода применяются при анализе наследования в популяции - для оценки концентрации мутаций и частот, мутирования. Эколого-физиологический метод оказывается необходимым для оценки действия абиотических и биотических факторов при определении приспособительной ценности фенотипов, относящихся к генетически различным классам особей. При этом большие экспериментальные возможности открывает моделирование действия отбора в искусственно создаваемых синтетических популяциях с заранее заданными генетическими параметрами - введение в популяцию тех или иных мутаций, инверсий, транслокаций и т. д.
Математический метод позволяет давать строгое количественное описание биологических процессов. Использование электронно-вычислительных машин явилось особенно перспективным для моделирования динамики генетической структуры популяции с учетом сложного взаимодействия многих факторов. Описываемые при этом ситуации в значительной мере приближаются к отражению истинной, сложной и противоречивой картины эволюционных процессов, протекающих в естественных популяциях растений и животных.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .


